前期白亜紀
前期白亜紀(ぜんきはくあき、Early Cretaceous)は、中生代の白亜紀を二分したうちのより古い方の地質時代。約1億4500万年前から約1億50万年前にあたり、古い順からベリアシアン期・バランギニアン期・オーテリビアン期・バレミアン期・アプチアン期・アルビアン期に分けられる[1]。
地理
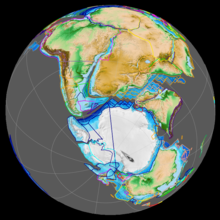
前期白亜紀の間にパンゲア大陸は分裂した。オーストラリア大陸と南極大陸(当時はまだ一つの地塊であった)およびインド亜大陸はゴンドワナ大陸から分離し、アフリカ大陸と南アメリカ大陸も分裂して初期の大西洋が形成され始めた。さらに先述の海進の影響で北アメリカ大陸は西側のララミディア大陸と東側のアパラチア大陸に分かれ、西部内陸海路が形成された。ヨーロッパは東ローラシア大陸との陸橋を形成し一部が陸続きとなった[2]。
現在のアメリカ合衆国ユタ州西部には山脈が形成され始め、その東側で降水量が増し湿潤化した。山脈を流れる河川により運搬された土砂は東側の平原に堆積し、カナダの北東部や南部に広がる広大な層準が形成された。この層準からはアラモサウルスの姉妹群であるモアボサウルスが発見されている[3]。
環境
温室効果ガスの放出により気候は現在よりも温暖な状態で安定しており[4]、南極大陸にも恐竜が生息できるほどであった[2]。ベリアシアン期からバレミアン期にかけては氷成堆積物が確認されていることから極域での氷床の存在が示唆されているが、アプチアン期とアルビアン期には存在しなかったと推測されている。またアプチアン期から後期白亜紀のコニアシアン期までは海水準が高く、多くの大陸の内陸部まで海進が起きていた[5]。
先述のアフリカ大陸と南アメリカ大陸の分裂により南大西洋が形成され始めたため、両大陸は湿潤化し、被子植物の多様化を可能とした[6]。現在のサハラ砂漠は乾燥帯ではなく、植物食動物の竜脚類や鳥脚類、およびそれを狙う獣脚類やワニ形上目が生息できるような豊かな土地であった[7]。
前期アプチアン期後期、末期アプチアン期から最初期アルビアン期、後期アルビアン期、末期アルビアン期にはそれぞれOAE1aからOAE1dまで計4回の海洋無酸素事変が起きた。原因としては、温暖化による海洋の成層構造の強化や、降水量・河川水量の増加による鉛直混合の弱化、一次生産の増大に伴う動物プランクトン増加に起因する酸素の大量消費などがあり、それぞれの無酸素事変の主要因については議論が続いている[5]。また、アプチアン期の最中である約1億2000万年前には南太平洋で二度に及ぶ大規模な火成活動が起き、莫大な玄武岩質溶岩が噴出して2000万立方キロメートルに達するオントン・ジャワ海台が形成された。この出来事はOAE1aの直前にあたり、噴火活動に起因する二酸化炭素をはじめとする温室効果ガスが先述の地球の温暖化と海洋無酸素事変をもたらしたとされる[8]。
生物
恐竜
前期白亜紀には前述の大陸移動により地理的隔離が起こり、恐竜の放散ならびに分化が進行した。特にジュラ紀に隆盛を誇った竜脚類は大きく様変わりすることになった。ディプロドクス上科はジュラ紀末から白亜紀最初期ごろに衰退し、ディクラエオサウルス科を北アフリカとアルゼンチンに残して姿を消した。またティタノサウルス形類を除く新竜脚類もジョバリアを残すのみになった。ティタノサウルス形類は衰退したディプロドクス上科やカマラサウルス科の生態的地位に入り込んで繁栄し、当時の南半球に位置したゴンドワナ大陸で繁栄した。彼らは南極大陸を介してオーストラリア大陸まで広がり、また当時アフリカ大陸と繋がっていたインド亜大陸にも進出した。また北アメリカ大陸とアジアにも進出し、鳥脚類が繁栄する中で一定の勢力を保っていた[2]。
獣脚類では多くのグループが白亜紀前期に出現しており、具体的にはオルニトミモサウルス類・テリジノサウルス上科・オヴィラプトロサウルス類・トロオドン科・ドロマエオサウルス科が姿を現した。また鳥類のうち真鳥類とエナンティオルニス類が出現したのも白亜紀前期である。ティラノサウルス上科は既に後期ジュラ紀の時点で出現していたものの、前期白亜紀時点ではディロングやエオティラヌスといった基盤的で小型の属のみに留まっていた[9]。なお、これら白亜紀前期の小型獣脚類には羽毛の存在が確認されているものも少なくない[10]。
鳥盤類の恐竜においては、鳥脚亜目のイグアノドンが4000万年以上に亘って繁栄した[11]。また、前期白亜紀末にはランジョウサウルスに代表される鳥脚亜目のハドロサウルス科が出現した。ハドロサウルス科は続く後期白亜紀に多様化を遂げ、他の鳥盤類恐竜と共に大部分の竜脚類の生態的地位を引き継ぐことになる[12]。また装盾亜目からは曲竜類(鎧竜)が出現しており、イギリスでヒラエオサウルスが下部白亜系から産出している[11]。一方で剣竜類は多様性が著しく低下した[4]。
恐竜以外の動物
海生爬虫類の魚竜は前期白亜紀の環境変動を受けて絶滅の一途を辿ることになった。先述の気温と海水準の上昇は魚竜の獲物・競合相手・生息域に影響を与え、魚竜に対して不利に働いた。魚竜の絶滅自体は約9000万年前にあたる後期白亜紀のセノマニアン - チューロニアン境界ごろのことであったが、魚竜の絶滅は二段階の過程を踏んでおり、多様性が低下したのは約1億年前のことであった[13][14][15]。ワニの仲間では、新鰐類のうち海棲適応を遂げたタラットスクス類が前期白亜紀を最後に絶滅した[16]。この他には新鰐類のテチスクス類(英語版)に属するサルコスクスがおり、このような巨大化を遂げたワニ形上目が恐竜と共存していた[7]。また、新鰐類の中からベルニサルティアやさらに派生的な正鰐類が出現した。正鰐類は現代型のワニ形上目のグループで、現生のワニも属する分類群である[17]。
哺乳類では約1億2500万年前にあたる中国の熱河層群義県層からエオマイアが発見されている。エオマイアは臼歯が3対あることから極めて初期の原始真獣類と判断されており、最古の有胎盤類の可能性もある。また前期白亜紀には有袋類の姉妹群である北半球後獣類と、現生のカモノハシに代表される単孔類も出現した[18]。
植物
植物相はシダや、キカデオイデア(英語版)属・ソテツ属・針葉樹といった裸子植物が大部分を占めていた。後期白亜紀以降に支配的になる被子植物は多様化を始めたばかりであった[3]。被子植物の出現は遅く見積もって1億2000万年前とされる。多様な環境を持っていたゴンドワナ大陸西部の熱帯高地で出現していたと見られる被子植物は、大西洋形成による湿潤化によりアフリカ大陸の熱帯低地に分布域を広げ、火山島群により往来が可能となっていた南アメリカ大陸にも進出した。その後は熱帯から高緯度地域へ分布を拡大したと考えられている[6]。
出典
- ^ “INTERNATIONAL CHRONOSTRATIGRAPHIC CHART(国際年代層序表)”. 日本地質学会. 2021年3月10日閲覧。
- ^ a b c 『世界の巨大恐竜博2006 生命と環境─進化のふしぎ』第134-135頁
- ^ a b 『世界の巨大恐竜博2006 生命と環境─進化のふしぎ』第140頁
- ^ a b デイヴィッド・E・ファストヴスキー、デイヴィッド・B・ウェイシャンペル 著、藤原慎一・松本涼子 訳『恐竜学入門 ─かたち・生態・絶滅─』真鍋真監訳、東京化学同人、2015年1月30日、271頁。ISBN 978-4-8079-0856-1。
- ^ a b 平野弘道、安藤寿男「白亜紀海洋無酸素事変」『石油技術協会誌』第71巻第3号、2006年、305-315頁、doi:10.3720/japt.71.305、ISSN 1881-4131。

- ^ a b 戸部博「花を咲かせる植物の辿った道」『生命誌』第18巻、JT生命誌研究館、1997年。
- ^ a b 小林快次『ワニと恐竜の共存 巨大ワニと恐竜の世界』北海道大学出版会、2013年7月25日、69-74頁。ISBN 978-4-8329-1398-1。
- ^ 『オントンジャワ海台における地球史上最大の火山活動の原因を解明』(プレスリリース)海洋研究開発機構、2012年2月17日。https://www.jamstec.go.jp/j/about/press_release/20120217_2/。2021年3月10日閲覧。
- ^ 『世界の大恐竜博2006 生命と環境─進化のふしぎ』第147頁
- ^ “刷新される恐竜像と私たち~21世紀の恐竜番組~”. NHK. 2021年3月10日閲覧。
- ^ a b ポール・バレット『ビジュアル百科 恐竜の生態 太古の王者はどう生きていたか』日経ナショナルジオグラフィック社、2006年7月13日。ISBN 4-931450-65-2。
- ^ 『世界の大恐竜博2006 生命と環境─進化のふしぎ』第150頁
- ^ 「魚竜の絶滅、気候変動が原因か 研究」『AFP BB News』(フランス通信)、2016年3月9日。2021年3月10日閲覧。
- ^ “【化石】イルカ似の爬虫類を絶滅に追い込んだ気候変動”. Nature Japan (2016年3月9日). 2021年3月10日閲覧。
- ^ Valentin Fischer; Nathalie Bardet; Roger B. J. Benson; Maxim S. Arkhangelsky; Matt Friedman (2016). “Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility”. Nature 7. https://www.nature.com/articles/ncomms10825.

- ^ 小林快次『ワニと恐竜の共存 巨大ワニと恐竜の世界』北海道大学出版会、2013年7月25日、22頁。ISBN 978-4-8329-1398-1。
- ^ 小林快次『ワニと恐竜の共存 巨大ワニと恐竜の世界』北海道大学出版会、2013年7月25日、28-31頁。ISBN 978-4-8329-1398-1。
- ^ 『世界の大恐竜博2006 生命と環境─進化のふしぎ』第122-123頁

参考文献
- 『世界の巨大恐竜博2006 生命と環境─進化のふしぎ』日本経済新聞社、NHK、NHKプロモーション、日経ナショナルジオグラフィック社、2006年。








